 Diplodocus (left) is a well-known sister taxon to the more robust and heavier Apatosaurus (right). In 1999, Mike Parrish and I published a study on the flexibility of the necks of these two diplodocids. Diplodocus, despite having a longer, more slender neck, is not as flexible dorsally as Apatosaurus. While both have 15 cervical vertebrae, those of Diplodocus are longer and much narrower, and significantly, the articular facets called zygapophyses (which guide and delimit flexibility between pairs of vertebrae) are smaller, particularly those
at the base of the neck, allowing Diplodocus substantially less ability to elevate its neck.
Of the two, Apatosaurus seemed more the generalist, while Diplodocus seemed highly specialized in its body plan, long neck, goofy teeth, and etc. It was naturally selected, therefore, to be one of the stars in the 1999 BBC series "Walking with Dinosaurs".
Diplodocus (left) is a well-known sister taxon to the more robust and heavier Apatosaurus (right). In 1999, Mike Parrish and I published a study on the flexibility of the necks of these two diplodocids. Diplodocus, despite having a longer, more slender neck, is not as flexible dorsally as Apatosaurus. While both have 15 cervical vertebrae, those of Diplodocus are longer and much narrower, and significantly, the articular facets called zygapophyses (which guide and delimit flexibility between pairs of vertebrae) are smaller, particularly those
at the base of the neck, allowing Diplodocus substantially less ability to elevate its neck.
Of the two, Apatosaurus seemed more the generalist, while Diplodocus seemed highly specialized in its body plan, long neck, goofy teeth, and etc. It was naturally selected, therefore, to be one of the stars in the 1999 BBC series "Walking with Dinosaurs".
| As the consultant for Episode 2 of the BBC series "Walking with Dinosaurs", when I arrived on location at Humboldt State Park (where the forest sequences were being filmed), series producer Tim Haines (below) immediately presented me with a problem: Diplodocus wasn't eating its conifers. Contrary to expectations, the needles just slipped between the peg-like teeth while larger branches just snagged on the tooth row. Without chisel-like dentition to chop up the tough conifer branches, it seemed improbable that these splayed teeth could have reduced the sticks of wood into a bolus to then swallow. | |
|
|
| So we changed its diet to horsetails and ferns, and the script to having the conifers just obstacles in the way of the sauropods in their search of this low browse. The peg-like dentition served well as rakes to gather up bite-sized mouthfuls. The tough woody conifer branches were left for those taller sauropods with chisel-like choppers such as Camarasaurus, Brachiosaurus. (Place the cursor over the scrolling images to momentarily pause the scrolling.) |
What limits intervertebral joint flexibility?
During intervertebral joint flexion, two pairs of pre- and postzygapophyses slide one across the other. Each pair is contained within a synovial capsule of tough ligaments. As flexion increses, eventually the surrounding capsule draws taut before they disarticulate. Our dissections suggested a "safety factor" of roughly 50% overlap. We learned subsequently that some vertebrates reduce the degree of zygapophyseal overlap at the limits of intervertebral flexion). Regardless the exact criterion (which is likely not constant across extant vertebrates) it was gratifying to hear Matthew Wedel and R. Kent Sanders say (at the 1999 SVP poster session) "we tried to prove you guys wrong, but couldn't". So our idea that zygapophyses geometrically limit the range of motion received independent "wet science" confirmation and seemed valid. They had x-rayed an ostrich neck at the limits of flexion, and further noted that "bone would break before the capsules would". The "safety factor" of overlap at the limit was likely less than 50%, but still substantial.
Sauropod neck vertebrae are opisthocoelous. The central articulations are ball-and-socket joints, with an anterior ball (condyle) on one fitting into deeply into a posterior socket (cotyle) on the next. The conformal articular surfaces in opisthocoely (and likewise heterocoely in avians) permits locating the instantaneous center of rotation from their geometry. Further geometric criteria regarding the limits of travel of the zygapophyseal facets permits estimating the flexibility of each such joint (and hence the flexibility of an entire neck).
(incidentally, some vertebrates such as bunnies have amphiplatyan central articulations with relatively thick intervertebral disks, and it's far more difficult to locate the instantaneous center of rotation, making estimation of range of motion far more difficult from osteology alone. Such is not the case given opisthocoely or heterocoely.)
There has been some disagreement regarding the "safety factor" of zygapophyseal overlap that remains when the living animal has reached its behavioral limit of flexion (laterally, or dorsally, etc.). Assuredly, vertebrates take care to not voluntarily disarticulate their (own) vertebral joints, hence some have osteological stops where the potential for injury is great (see the discussions regarding camel and giraffe, below). Such osteological bracing is not universal however. It not present in the canid neck, for example. So what keeps a dog's neck from disarticulation? Ultimately, the ligaments of the articular capsules, and before that, the strength of its overlying musculature. The soft tissues that protect the neck also bind it before the articulation reaches is minimum degree of overlap. That theoretical minimal overlap might be substantially less than 50% in some vertebrates, but it is certainly greater than zero. And in vivo, muscles, ligaments, and fascia often
have further limited movement, thus the digital manipulations we reported represent what we believe is a "best case" scenario for their neck mobility.
Some vertebrates have osteological stops which prevent the zygapophyses from disarticulating. Camels (right) and horses, for instance, have pits that limit the dorsiflexion of the more cranial vertebra's postzygapophyses. The rhino is massively braced against disarticulation in lateral flexion as well; the neck becomes rigid at the limits, which is of clear advantage to an animal engaged in its particular lifestyle. Interestingly, the giraffe's neck is also stabilized against disarticulation in lateral flexion by osteological bracing (see below).
|
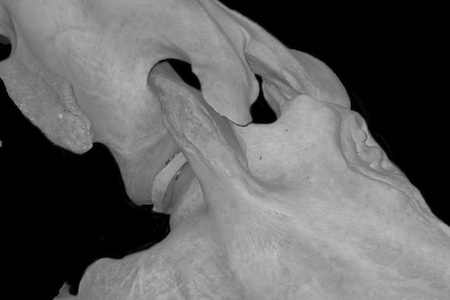 |
The camel (right) is capable of dorsiflexing its neck at the base such that it can rub its occiput against its shoulders (it's a male thing, apparently). While at an SVP meeting the inset picture (from Gauthier-Pilters and Daag, 1981) was offered to be proof positive that zygapophyses can be disarticulated, the camel's neck is actually comfortably braced and in articulation, despite appearances. Two further points to make here: First, sauropod necks lack such osteological bracing and hence are often found dorsiflected into an extreme "death pose"; wherein the zygapophyses are completely disarticulated (examine the death pose for Camarasaurus and Euhelopus). Secondly, it has been suggested (and I'm not making this up) that all (!) tetrapods habitually maximally dorsiflex at the base of the neck when alert. Here is what "maximal dorsiflexion " means for a camel. A camel is no where near the limits of dorsiflexion when merely alert. |
 |
| The giraffe neck (like many extant vertebrates) has osteological stops at the base of the neck to prevent disarticulation in ventriflexion and dorsiflexion, and lateral bracing to preserve articulation of the zygapophyses. The total mobility of the giraffe's neck can be replicated rather well by assembling a digital representation of the cervical series (in this case, based on CT data courtesy the American Museum of Natural History), then after carefully establishing the centers of rotation for each intervertebral joint, exercising them together until their osteological stops are encountered. At the limits of travel the zygapophyses in fact preserve substantial overlap. The giraffe neck thus becomes increasingly stabilized at the limits, particularly laterally, and primarily at the base of the neck where the potential for injury is greatest. Lateral bracing is present in some sauropods such as Diplodocus, permitting a stable yet light vertebral design that allowed even greater length (in Barosaurus), in contrast to Camarasaurus and Apatosaurus which rely on massive, flat zygapophyses and wide lateral processes to resist such torsional loading. |  |
Recently I have created animations where, for example, Apatosaurus or Camarasaurus is given the exercise of creating a reflex curve so that it can explore not just the extreme outer surface of its "feeding envelope" but also reach back to explore its "reachability volume". This work, first shown at the 2010 SVPCA in Cambridge show that sauropods are miserable at this task which is so easily achieved by many long-necked birds. Sauropods can sweep back and forth and up and down very well, but they are barely able pull the head in and out radially to develop that two-dimensional feeding surface into a three-dimensional feeding volume. These long necks are better-suited for broad swath-like feeding patterns, not selective feeding.
So what is the general conclusion for Diplodocus? Dentition that are great for soft low browse, a neck that sweeps laterally and downward much more than upward, a neck that doesn't really bend as much as the rubber toys would suggest. And relative to Apatosaurus, for Diplodocus (and even more so than Barosaurus) the zygaophyses at the base of the neck are so slender they permit much less dorsiflexion than Apatosaurus. Whatever the precise limit on the range of motion at each intervertebral joint, if that posited criterion is applied equally to these two diplodocids, Apatosaurus shows significantly more flexibility than Diplodocus, particularly in dorsiflexion, despite the longer neck.
That Diplodocus had less dorsal flexibility than
Apatosaurus has been trivialized into our supposedly suggesting that "diplodocids could not raise their necks far above the horizontal". But in the 1999 study we stated "The posture for the trunk and limbs was based on previous reconstructions and not addressed by this study". Despite the downsloping backs of Hatcher's and Gilmore's early reconstructions, we estimated the head could be raised to over 4 m for Diplodocus and 6 m for Apatosaurus. Needless to say, that is "far above the horizontal". Subsequently, when we revised the sauropod body plan to incorporate current notions of pectoral girdle placement, which brought the anterior dorsal column to roughly horizontal, the estimates of maximum head heights increased accordingly. On many occasions since 1999 we have presented and published visualizations with the neck associated with revised diplodocid body plans (see below)
These revised models are shown in various presentations, including:
- 2010 SVPCA (Cambridge)
- 2009 SVP ( Bristol)
- 2005 German Sauropod Group (Aathal)
- 2002 SVP (Norman)
- 2001 SVP (Bozeman)
and publications:
- Stevens, Kent A. and J. Michael Parrish (2005) Neck posture, dentition and feeding strategies in Jurassic sauropod dinosaurs. In Thunderlizards: the sauropodomorph dinosaurs. Virginia Tidwell and Kenneth Carpenter, eds. Bloomington: Indiana University Press.
- Stevens, Kent A. and J. Michael Parrish (2005) Digital reconstructions of sauropod dinosaurs and implications for feeding. The Sauropods: Evolution and paleobiology. Kristi Curry Rogers, and Jeffrey Wilson, eds. Berkeley: University of California Press.
So how did Diplodocus hold its head? Out in front, or raised up?
Since the late 1990s the primary emphasis of my research (regarding sauropods at least), has been to try to sort out what the osteology says about the axial skeleton of sauropods. Starting with diplodocids then Camarasaurus then various other sauropods, we found that all their necks were straight extensions of the anterior dorsal vertebral columns. No evidence for a giraffe-like upward bend in the neck. Just straight off the shoulder. That met with little resistance for diplodocids, which have long been expected to have been low browsers by all but a few cartoonists, manufacturers of rubber dinosaur toys, and the odd British museum curator or two. Regarding other sauropods such as Camarasaurus and Brachiosaurus, the swan-neck interpretation of the rubber toys has been harder for some to accept. There is apparently growing acceptance of the fact that sauropod neck osteology does not provide an intrinsic upward bend to the base of the neck as it does in the giraffe and many birds.
But that's just the osteology: the necks fit together as straight extensions of the back as it emerges from the shoulder. Sauropod necks of all sorts are pretty straight in the middle, and bit droopy out near the head, when the bones are all assembled. But what about how they were held when they were alive? Well, that's perhaps another story.