Note: The work originally described below is leading to a new animation of the process by which T. rex might have sat and stood. When available this page will be revised.
For the June, 2005 Symposium "100 Years of Tyrannosaurus rex" hosted by the Black Hills Institute in Hill City, SD, I presented a paper "Rex, sit". The challenge was to try to understand how Tyrannosaurus rex might have transitioned between a bipedal, standing posture and a resting pose. It has long been expected that the great theropod would have settled its weight upon its pubic boot. Just how might it have sat down and gotten up again? this question was explored using an articulated DinoMorph™ model built in collaboration with Art Andersen of Virtual Surfaces, Inc. and the Black Hills Institute of Geologic Research.

Ray Wilhite using an Immersion Microscribe® digitizer on casts of elements of the pelvic girdle of Tyrannosaurus rex specimen ‘Stan’ BHI-3033 (photo courtesy Ray Wilhite and Virtual Surfaces, Inc.).

Completed DinoMorph™ model of Tyrannosaurus rex specimen BHI-3033‘Stan’. The appendicular skeleton and head were digitized; the remainder of the axial skeleton was represented in a more schematic form, with important parameters (e.g., centrum length, neural spine height, and intervertebral separations) accurate based on measurements taken off of the specimen. Since 2005, I have completely revised the model, as you can see here.

The axial skeleton was laid out with reference to measurements taken from the mount and photographs of BHI-3033 'Stan'. Scale bar indicates 11.2 m overall length.
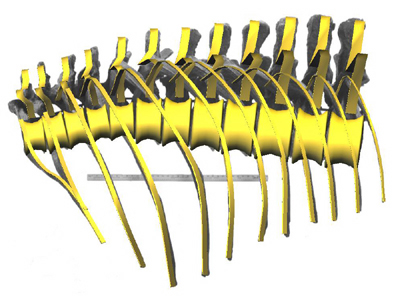
The reconstruction of the trunk superimposed upon a reference photograph of an assembly of casts of the ‘Stan’ specimen. Background image courtesy Black Hills Institute of Geologic Research (yardstick for scale).
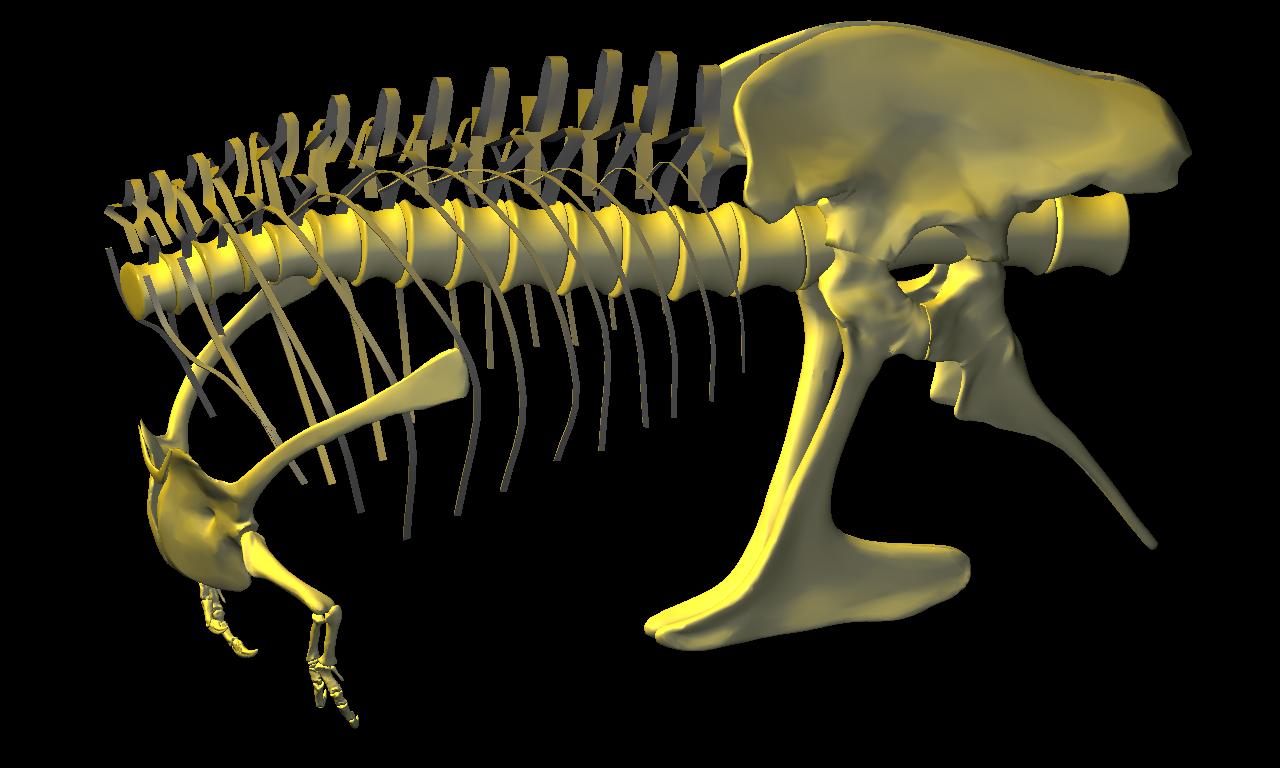
DinoMorph™ model shown with addition of digitized pectoral girdles (including furcula), forelimbs and pelvic girdle, all based on ‘Stan’.

Visualization of the distribution of body mass, based on a parametric fit to segments of body cross-section of the axial and appendicular skeleton.
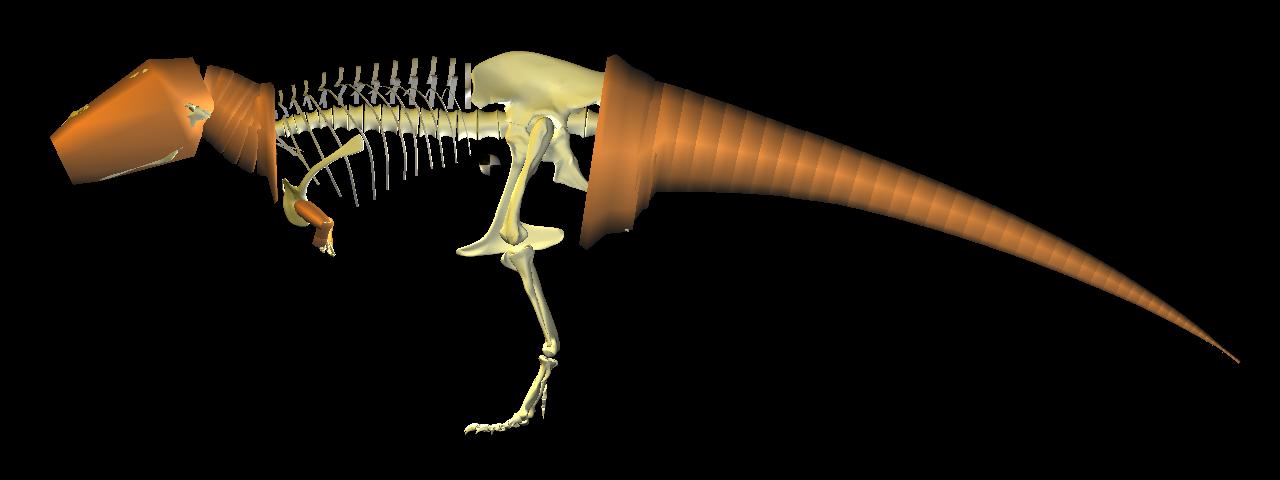
Computed center of mass visualized just anterior pubis, and above pes, as required for static bipedal balance.
 |
 |
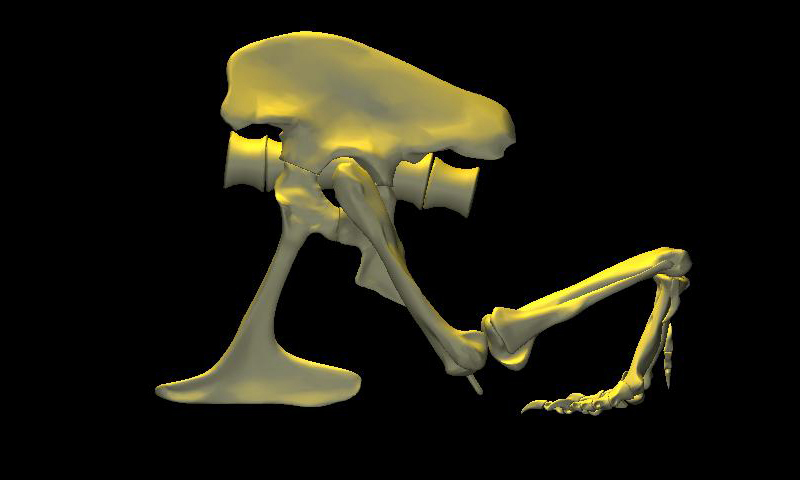 |
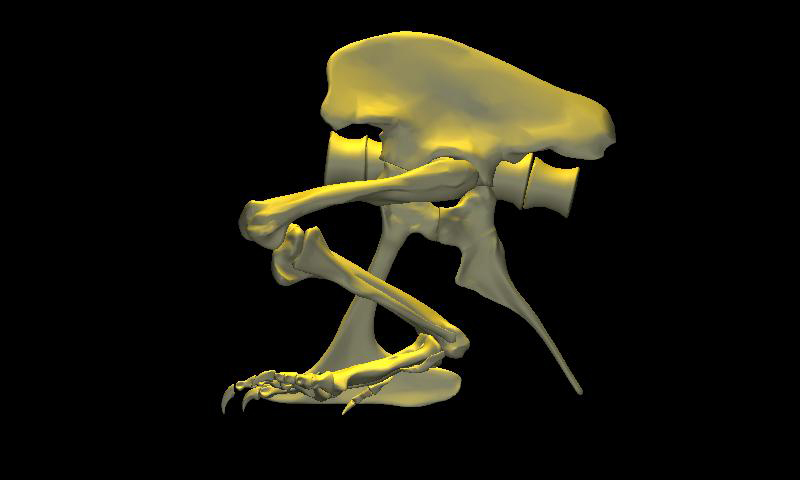
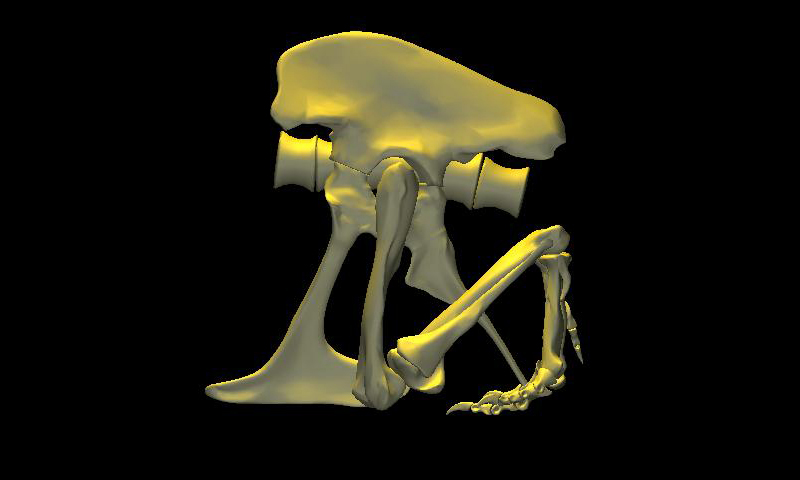
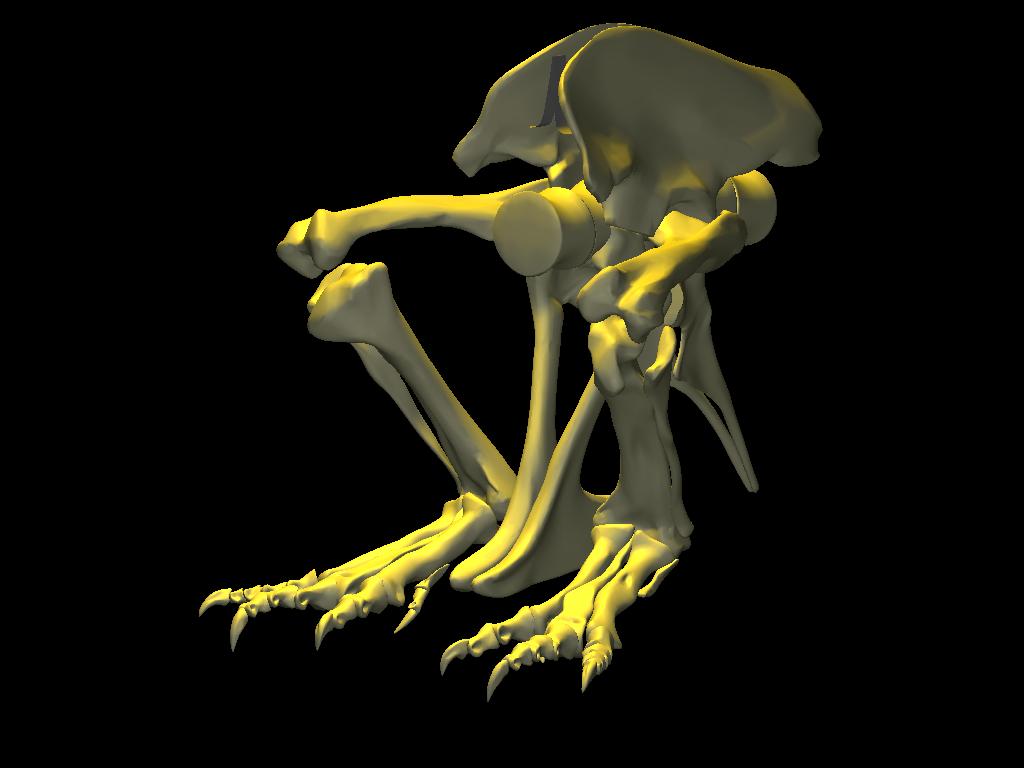
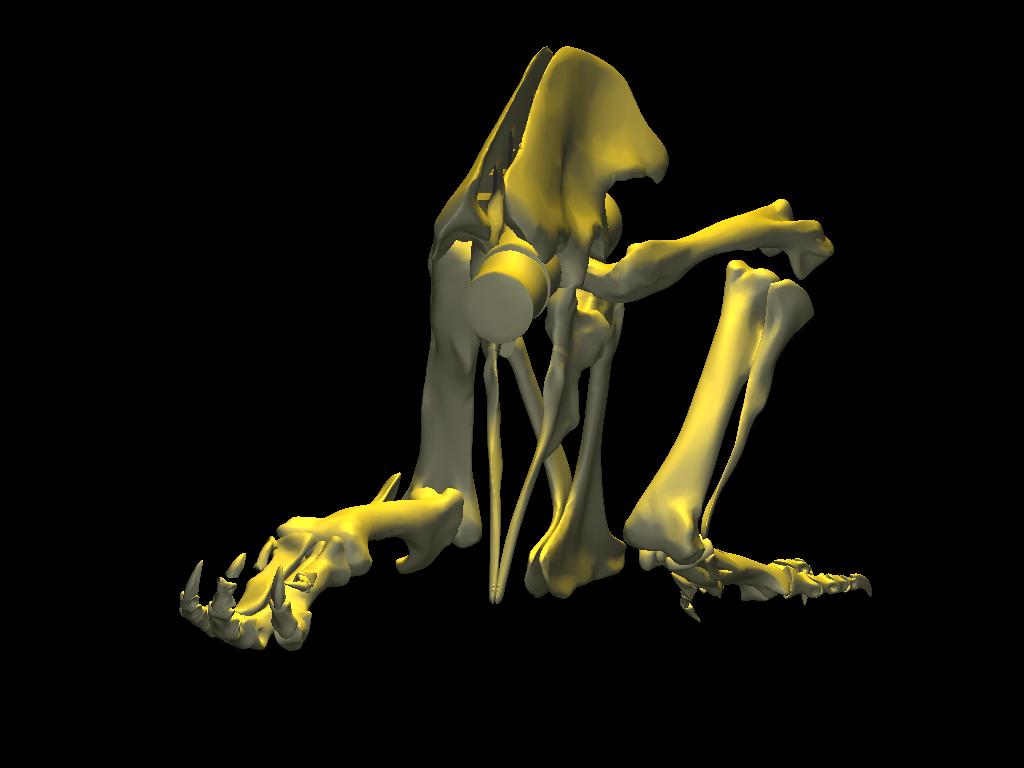
DinoMorph™ model of T. rex ‘Stan’ demonstrating that the pubic shaft is sufficiently long to allow the hindlimb to assume a broad range of positions from crouch, to kneel to moderate extension of the hip and knee, to full leg extension when stretched behind the hips (not shown), all with the animal’s weight resting upon pubic boot. Note that in these three images the tarsus remains posed at near the limit of hyperflexion; in the left two images the knee is fully flexed. Observe that the knee just clears the ground as it swings through hip flexion/extension. Joint flexibility estimated in collaboration with Yoshio Ito and Kenneth Carpenter.
 |
 |
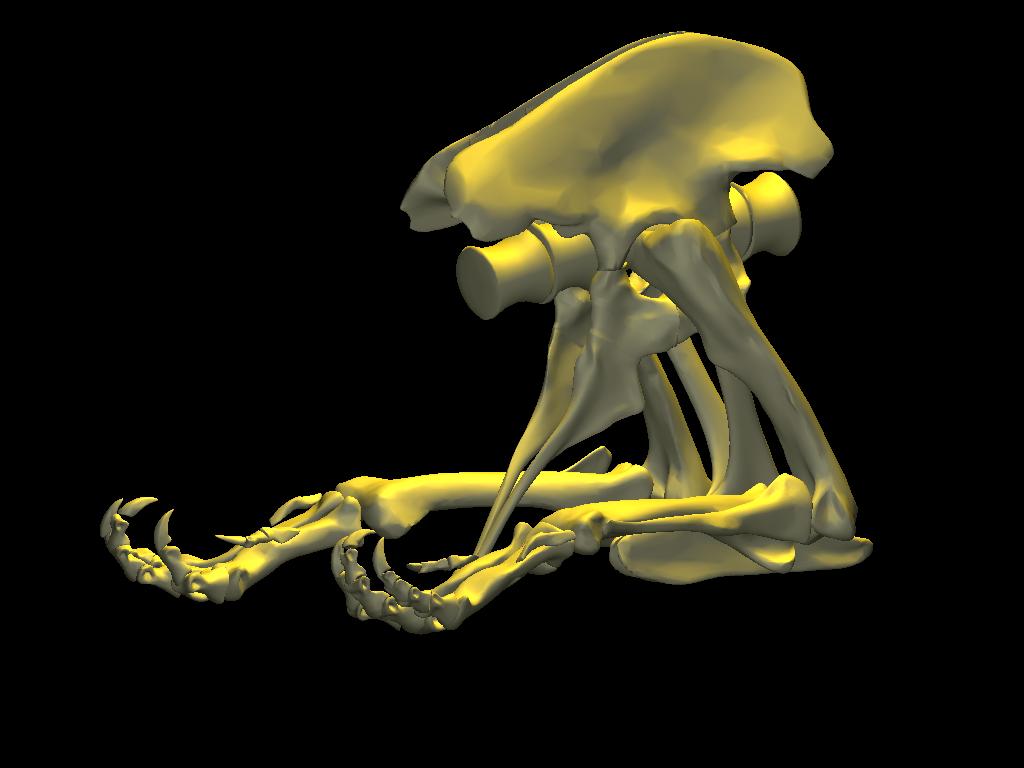 |
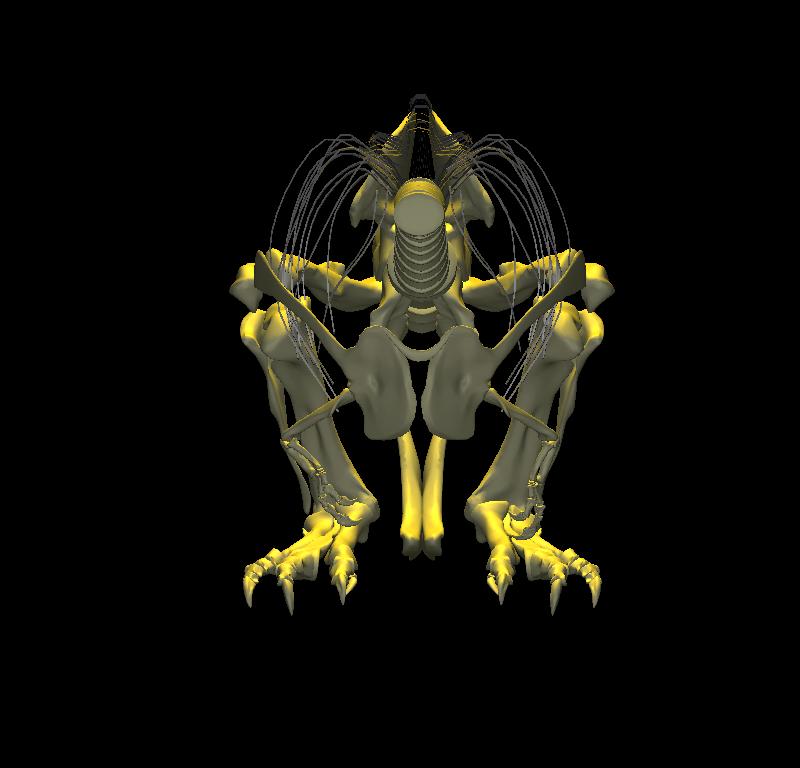
With the body mass supported by the pubic boot, the hind limbs appear to have been able to shift from a sitting position (with hip flexed) to kneeling on one knee or both knees, without having to lift the body weight off of the pubic boot. While the axis-like insertion of femur head within acetabulum suggest little femoral abduction was possible, there was likely sufficient flexibility to provide lateral stability. Moreover, the posterolateral angulation of the acetabular axis caused the knees to splay with femoral protracted, again aiding stability against lateral tipping in addition to clearing the ribcage as necessary in locomotion.
 |
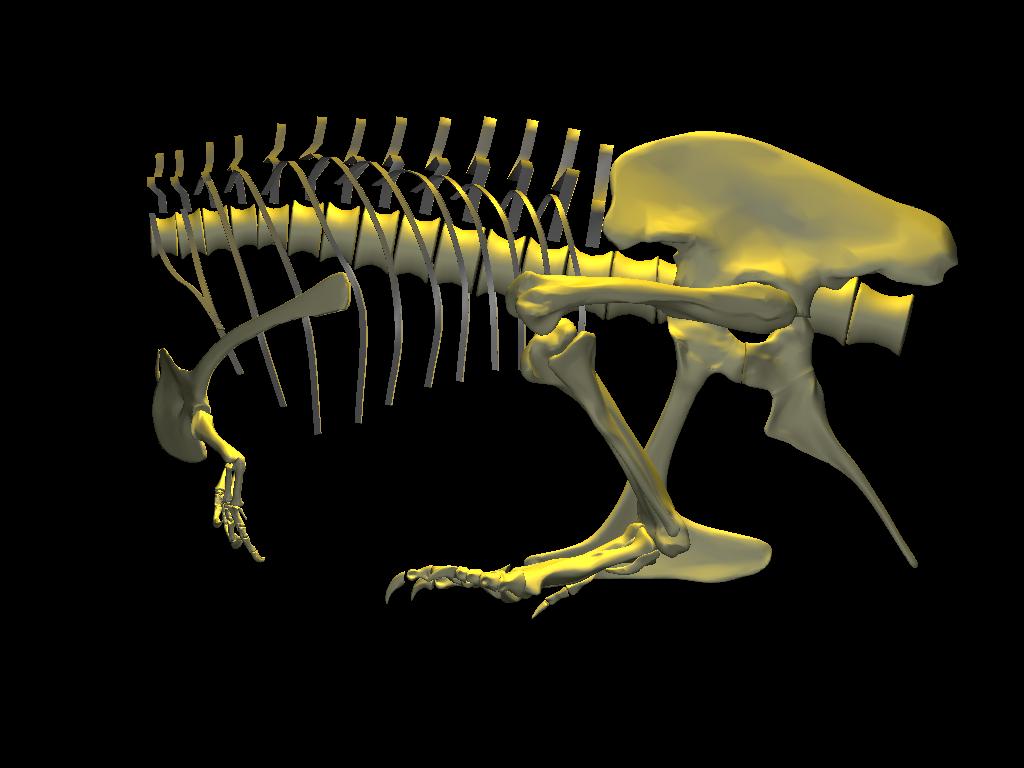 |
When resting upon the pubis, the forelimbs are near ground level. They are brought closer to ground level as a consequence of initiating a standing movement from this resting pose. A modest tipping of the body, by pivoting about the curved anteroventral surface of the prepubis, would have shifted the overall center of mass anteriorly and returned the point of balance to between the hindfeet. In the process, the forelimbs would have been available to assist in stabilizing (if not actively contributing towards elevating) the body by pushing against the ground.
=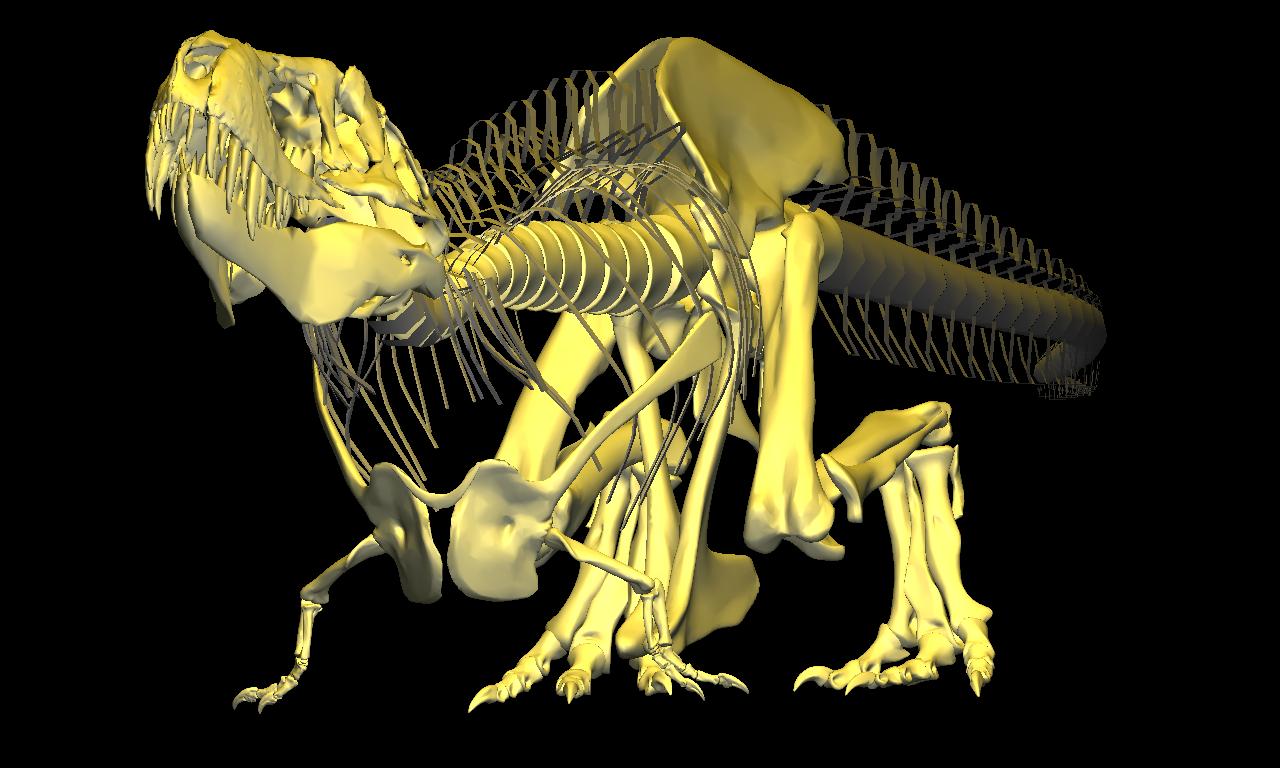
=Reconstruction of the pectoral girdles and forelimbs based on CT data (but for the radiale and distal carpal, which were sculpted within DinoMorph™). Note that with elbows and manus extended, the forelimbs can act as a ‘jack stand’ to stabilize the body during ascent, but that the line of action of the ground reaction force would have placed the furcula under significant bending stress, consistent with commonly observed healed fractures. Forelimb range of motion estimated in collaboration with Kenneth Carpenter.
Here are movies showing possible descent (.mov format 3.9 MB) and ascent ( .mov format, 3.7 MB) movements.